


学習目標
- ニューロン(神経細胞)の構造的構成要素の機能を挙げ、説明する
- ニューロンの4つの主要なタイプを挙げ、説明する
- 異なる種類のグリア細胞の機能を比較する
動物の神経系
動物界の神経系は、図35.2に示したさまざまな動物が示すように、その構造や複雑さに違いがあります。海綿動物のように、真の神経系を持たない生物もいます。また、クラゲのように、脳を持たず、代わりに “神経網 “と呼ばれるニューロン(神経細胞)が別々につながっているシステムを持つものもいます。ウミウシなどの棘皮動物は、神経細胞が束ねられて神経と呼ばれる繊維になっています。扁形動物門の扁形動物は、小さな「脳」と2本の神経索からなる中枢神経系(CNS)と、全身に伸びる神経系を含む末梢神経系(PNS)の両方を持っています。昆虫の神経系はより複雑ですが、かなり分散化されており、脳、腹神経索、神経節(神経細胞がつながったクラスター)があります。これらの神経節は、脳からの入力なしに運動や行動を制御することができます。タコは、無脊椎動物の神経系の中で最も複雑な神経系を持っているかもしれません。タコは、脊椎動物の種と構造的に類似した特殊な葉と目に組織化されたニューロンを持っています。
脊椎動物の神経系は、無脊椎動物に比べ、より複雑で、中枢性、特殊性に富んでいます。脊椎動物の神経系は、脳と脊髄からなる中枢神経系(CNS)と、末梢の感覚神経や運動神経からなる末梢神経系(PNS)という基本的な構造は共通しており、それぞれに大きな多様性があります。無脊椎動物と脊椎動物の神経系の興味深い違いの一つは、多くの無脊椎動物の神経索が腹側にあるのに対し、脊椎動物の脊髄は背側にあることです。進化生物学者の間では、これらの異なる神経系計画が別々に進化したのか、それとも脊椎動物の進化の過程で無脊椎動物の身体計画の配置が何らかの形で「反転」したのか、議論がなされています。
学習のためのリンク
ハーバード大学の生物学者Mark Kirschnerが、脊椎動物の進化における「反転」現象について語るビデオをご覧ください。
神経系
神経系は、化学的・電気的信号を受信・伝達する特殊な細胞であるニューロン(神経細胞)と、ニューロンと補完的に情報処理を行い、ニューロンのサポート機能を担う細胞であるグリア細胞から構成されています。
ニューロンは電線に例えることができ、ある場所から別の場所へ信号を伝達します。グリア細胞は、電線が正しい場所に届くようにしたり、電線をメンテナンスしたり、切れた電線を取り除いたりしている電力会社の社員に当たります。グリア細胞は社員に例えられるものの、最近の証拠によれば、グリア細胞もまた、神経細胞のシグナル伝達機能の一部を利用していることが示唆されています。
神経系の各部位に存在するニューロンとグリア細胞の種類には、非常に多様性があります。ニューロンは大きく分けて4種類あり、いくつかの重要な細胞成分を共有しています。
ニューロン Neurons
実験用のショウジョウバエDrosophila melanogasterの神経系には、ロブスターと同じ数の約10万個のニューロンがあります。これに対し、マウスは7,500万個、タコは3億個です。そして人間の脳には約860億個のニューロン存在します。このように数は大きく異なるものの、こうした動物の神経系は、基本的な反射行動から、餌を探したり、相手に求愛したりするような複雑な行動まで、多くの同じ行動を制御しています。こうした行動の根底には、ニューロン同士、あるいは他の種類の細胞とのコミュニケーションの能力があります。
ほとんどのニューロンは、同じ構成要素を共有しています。しかし、ニューロンは高度に特殊化されており、異なる種類のニューロンは、その機能的役割に関連して異なるサイズと形状を有しています。
ニューロンを構成する部品 Parts of a Neuron
他の細胞と同様に、各ニューロンは、核、滑面小胞体と粗面小胞体、ゴルジ体(ゴルジ装置)、ミトコンドリア、および他の細胞構成要素を含む細胞体(またはソーマ)を持っています。また、ニューロンは、図35.3に示すように、ニューロンのコミュニケーションを可能にする電気信号を受信し、送信するための独自の構造を持っています。 樹状突起は樹木のような構造で、細胞本体から伸びて、シナプスと呼ばれる特殊な接合部で他のニューロンからのメッセージを受信します。樹状突起を持たないニューロンもありますが、複数の樹状突起を持つタイプのニューロンもあります。樹状突起は、樹状突起スパインと呼ばれる小さな突起を持つことがあり、これがシナプス結合しうる表面積をさらに増大させます。
樹状突起で受信された信号は、細胞体へと受動的に移動します。細胞体には、複数のシナプスからの信号を統合する軸索小丘という特殊な構造があり、細胞体と軸索の接合部として機能します。軸索はチューブ状の構造で、統合されたシグナルを軸索末端と呼ばれる特殊な末端に伝達します。軸索末端は、他のニューロン、筋肉、または標的器官とシナプスを形成します。軸索末端で放出される化学物質によって、信号はこれらの他の細胞に伝達されます。
ニューロンは通常、1~2本の軸索を持ちますが、網膜のアマクリン細胞のように軸索を持たないニューロンもあります。軸索の一部は髄鞘(ミエリン鞘)で覆われており、髄鞘が絶縁体となって軸索を伝わる電気信号の散逸を最小限に抑え、伝導速度を大幅に向上させます。ヒトの運動ニューロンの軸索は、背骨の付け根から足の指まで、1メートルもの長さがあるため、この絶縁性は重要です。髄鞘は、実はニューロンの一部ではありません。髄鞘はグリア細胞によって作られるのです。軸索に沿って、髄鞘には周期的な隙間が存在します。これらの隙間はランビエ絞輪と呼ばれ、信号が軸索に沿って移動する際に「充電」される場所です。
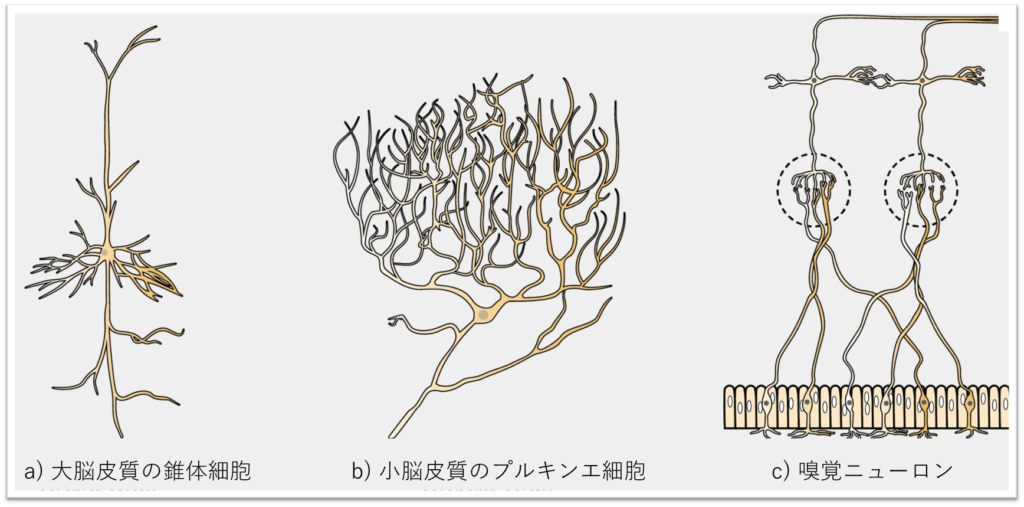
ニューロンの情報伝達は、ニューロン同士(および筋肉細胞などの他の細胞)の結合に依存しています。一つのニューロンからの樹状突起は、他の多くのニューロンからシナプスの接触を受けることがあります。例えば、小脳のプルキンエ細胞からの樹状突起は、20万もの他のニューロンから接触を受けていると考えられています。
VISUAL CONNECTION
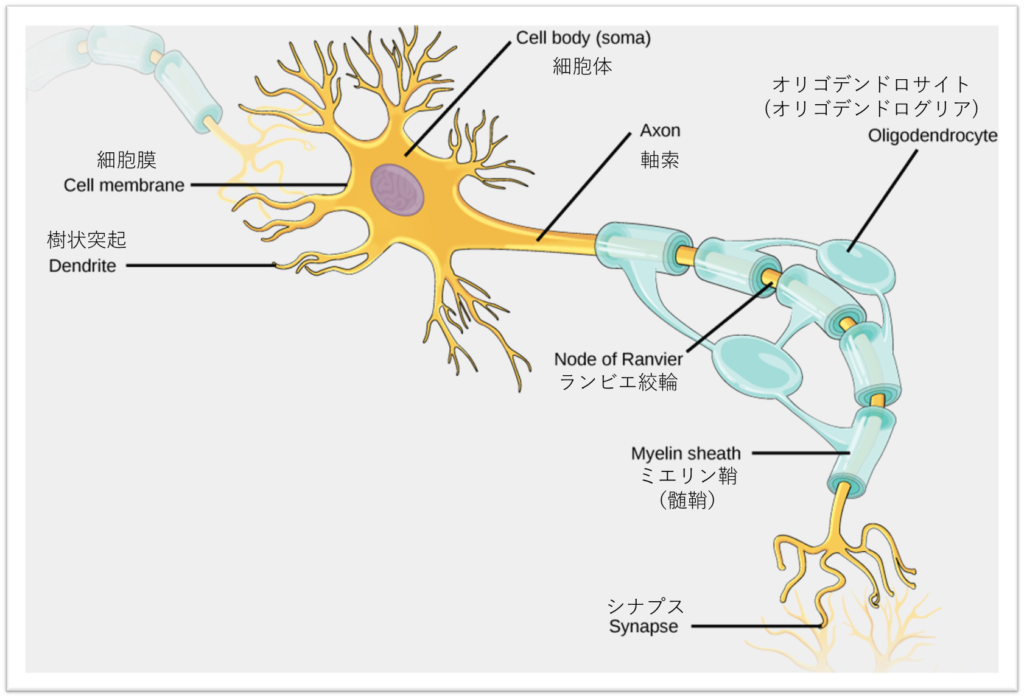
次の記述のうち、誤っているものはどれか。
- ソーマはニューロンの細胞体である。
- 髄鞘は、樹状突起に絶縁層を提供する。
- 軸索はニューロンから標的への信号を伝達する。
- 樹状突起は信号を細胞体へ伝える。
ニューロンの種類
ニューロンにはさまざまな種類があり、あるニューロンの機能的役割は、その構造に密接に依存します。図35.4に示されたニューロンからもわかるように、神経系のさまざまな部分で(および種を超えて)見られるニューロンの形や大きさは驚くほど多様です。
ニューロンには多くのサブタイプが定義されていますが、大きく分けて、単極性、双極性、多極性、および偽単極性の4つの基本型に分類されます。図35.5は、これらの4つの基本的なニューロンを示したものです。
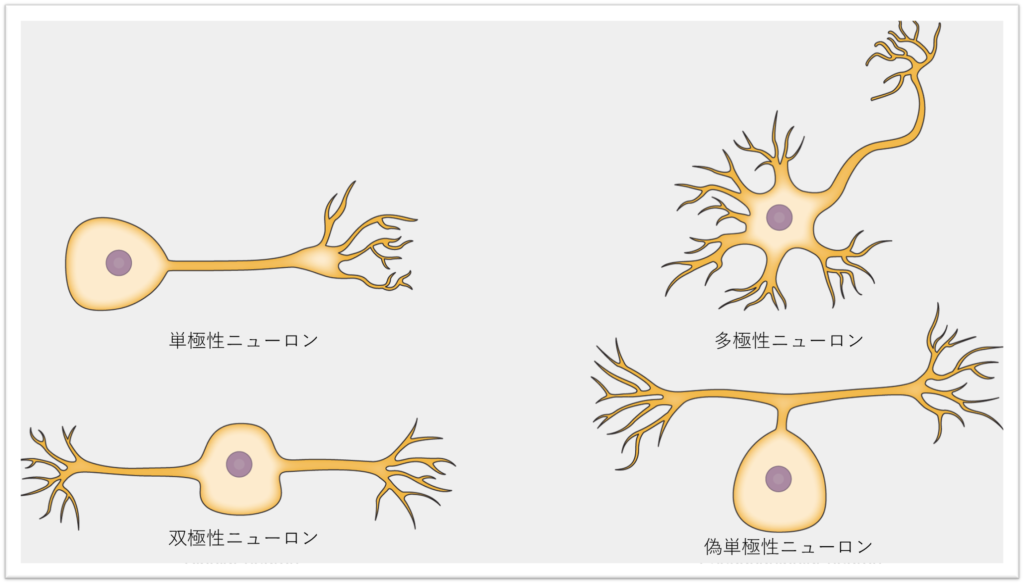
単極性ニューロンは、細胞体から離れる方向に伸びる1つの構造のみを持ちます。このニューロンは脊椎動物には存在しませんが、昆虫には存在し、筋肉や腺を刺激します。
双極性ニューロンは、細胞体から1本の軸索と1本の樹状突起が伸びています。例えば、網膜双極細胞は、光に敏感な視細胞から信号を受け取り、その信号を脳に伝える神経節細胞に伝達します。多極性ニューロンは、最も一般的なタイプのニューロンで、1本の軸索と複数の樹状突起を含みます。
多極性ニューロンは、中枢神経系(脳と脊髄)に見られます。多極性ニューロンの例は、小脳のプルキンエ細胞で、多くの枝分かれした樹状突起を持ちますが、軸索は1本だけです。
偽単極性ニューロンは、単極性と双極性の両方の特徴を備えています。偽単極性ニューロンは単極性ニューロンのように細胞体から1つの突起が伸びていますが、この突起は後に双極性ニューロンのように2つの異なる構造に分岐します。ほとんどの感覚ニューロンは偽単極性で、軸索は2つの延長に分かれ、1つは感覚情報を受け取る樹状突起に接続され、もう1つはこの情報を脊髄に伝達するものとなります。
EVERYDAY CONNECTION:ニューロン新生
かつて科学者たちは、人は生まれながらにしてすべてのニューロンを持っていると信じていました。しかし、ここ数十年の研究により、ニューロンの新生が大人になっても続いていることが明らかになりました。ニューロン新生は、歌を学習する際に新しいニューロンを生成する鳴禽類で初めて発見されました。哺乳類の場合、新しいニューロンは学習にも重要な役割を果たします。海馬(学習と記憶に関与する脳の構造)では、毎日約1000個の新しいニューロンが発生します。新生ニューロンのほとんどは死んでしまいますが、海馬で生き残っている新生ニューロンの数の増加は、ラットが新しい課題をどれだけうまく学習できたかと相関していることが、研究者らによって明らかにされました。興味深いことに、運動と一部の抗うつ剤も、海馬のニューロン新生を促進します。ストレスは逆効果です。ニューロン新生は、他の組織の再生に比べるとかなり限定的ですが、この分野の研究が、アルツハイマー病、脳卒中、てんかんなどの疾患の新しい治療法につながる可能性があります。
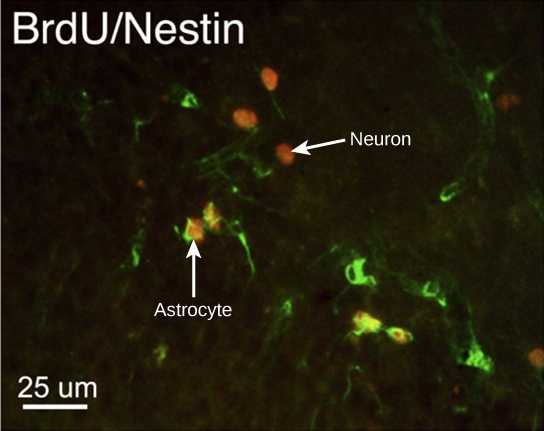
新しいニューロンはどのようにして発見されるのでしょうか?研究者は、ブロモデオキシウリジン(BrdU)と呼ばれる化合物を動物の脳に注射することができます。すべての細胞がBrdUに曝露されますが、BrdUが取り込まれるのは、S期にある新しく生成された細胞のDNAのみです。免疫組織化学と呼ばれる技術を使って、取り込まれたBrdUに蛍光標識を付けることができます。そして、研究者は蛍光顕微鏡を使って脳組織中のBrdU、つまり新しいニューロンの存在を視覚化することができるのです。
図35.6は、ラットの海馬に存在する蛍光標識されたニューロンを示す顕微鏡写真です。
グリア細胞
グリア細胞は神経系の脇役と思われがちですが、実は脳内のグリア細胞の数はニューロンの数を10倍も上回っています。ニューロンは、このグリア細胞の重要な役割なしには機能しません。グリア細胞は、発達中のニューロンを目的地に導き、ニューロンに害を与えるイオンや化学物質を緩衝し、軸索の周りに髄鞘を提供します。また、グリア細胞は神経活動に反応し、ニューロン間のコミュニケーションを調整する役割を担っていることも、最近になって発見されました。グリア細胞が正しく機能しないと、脳腫瘍の多くはグリア細胞の突然変異によって引き起こされることからも分かるように、悲惨な結果になることがあります。
グリア細胞の種類
グリア細胞には機能の異なるいくつかの種類があり、そのうちの2つを図35.7に示します。
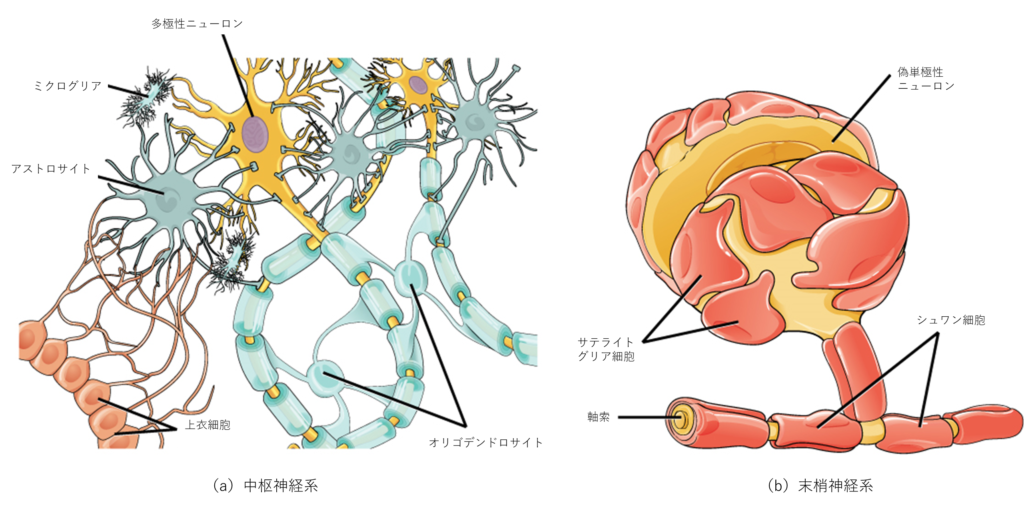
図35.8aに示すアストロサイトは、中枢神経系(CNS)内の毛細血管とニューロンの両方に接触しています。アストロサイトは、神経細胞に栄養やその他の物質を供給し、細胞外液中のイオンや化学物質の濃度を調節し、シナプスを構造的に支えます。また、アストロサイトは、血液脳関門(脳への有害物質の侵入を防ぐ構造)を形成しています。アストロサイトは、神経活動に応じて活性化し、アストロサイト間でカルシウム波を伝達し、周囲のシナプスの活動を調節することがカルシウムイメージング実験により明らかにされています。
サテライトグリア細胞は、末梢神経系(PNS)のニューロンに栄養を供給し、構造的なサポートを提供します。
ミクログリアは、死んだ細胞を除去し、分解し、侵入してきた微生物から脳を保護します。
図35.8bに示すオリゴデンドロサイトは、中枢神経系(CNS)の軸索の周囲に髄鞘を形成します。一つの軸索は数個のオリゴデンドロサイトによって髄鞘化(ミエリン化)され、一個のオリゴデンドロサイトは複数のニューロンに対して髄鞘を供給することが可能です。このことは、末梢神経系(PNS)と異なる点です(末梢神経系では、シュワン細胞全体が軸索を取り囲んでいるため、1つのシュワン細胞が1つの軸索にのみ髄鞘を供給します)。
放射状グリア細胞は、発達中のニューロンが最終目的地に移動する際の足場として機能します。
上衣細胞は、液体に満たされた脳の脳室と脊髄の中心管を覆っています。上衣細胞は、脳のクッションである脳脊髄液の産生や循環に関与するほか、脈絡叢の構成要素となっています。


コメント