


学習目標
- 現在の進化論がどのように発展してきたかを説明する
- 適応の定義
- 収斂進化と分岐進化を説明する
- 相同的構造と痕跡器官について説明する
- 進化論に関する誤解について話し合う
自然選択による進化は、時間の経過とともに種が変化するメカニズムを説明するものです。ダーウィンがこの考えを探求し始めるずっと以前から、科学者、哲学者、研究者などがこのテーマについて提案し、議論していました。古代ギリシャの哲学者プラトンは、種は静的で不変であることを著作で強調していますが、古代ギリシャ人には進化論的な考えを示す人もいました。18世紀には、博物学者のジョルジュ=ルイ・ルクレール・ド・ビュフォン Georges-Louis Leclerc Comte de Buffon が動物の進化に関する考えを再び導入し、環境が似ていても地域によって植物や動物の個体数が異なることを観察しています。また、この頃、絶滅した種が存在することを認める者もいました。
また、18世紀には、スコットランドの地質学者・博物学者であるジェームズ・ハットン James Hutton が、地質学的変化は、現在と同じように働くプロセスによる小さな変化が長い時間をかけて積み重なることで徐々に起こったという説を唱えました。これは、地球の地質が比較的短い期間に起きた大変動の結果であるという当時支配的だった考えと対照的でした。19世紀の地質学者チャールズ・ライエル Charles Lyell は、ハットンの考えを広めました。彼はダーウィンの友人でもあります。ライエルの考えは、ダーウィンの考え方に影響を与えました。地球の年齢が高いというライエルの考え方は、種が徐々に変化するための時間をより多く与え、その変化の過程を類推させるものだったのです。19世紀初頭、ジャン=バティスト・ラマルク Jean-Baptiste Lamarck は、進化的変化のメカニズムを詳細に記した本を出版しました。現在では、環境による個体の変化、あるいは子孫の生涯における構造の用・不用により、種に変化をもたらすとするこのメカニズムは「獲得形質の遺伝」と呼ばれます。多くがこの進化的変化のメカニズムを否定しましたが、ラマルクの考え方は進化論に重要な影響を与えました。
チャールズ・ダーウィンと自然選択
19世紀半ば、チャールズ・ダーウィンCharles Darwin と アルフレッド・ラッセル・ウォレス Alfred Russel Wallaceという2人の博物学者が、それぞれ独自に実際に起きている進化のメカニズムを考え、記述しました。重要なことは、それぞれの博物学者が熱帯地方への遠征で自然界の探索に時間を費やしたことです。ダーウィンは、1831年から1836年にかけて、ビーグル号で南米、オーストラリア、アフリカ大陸南端を含む世界一周をしました。ウォレスは、1848年から1852年までブラジルのアマゾン熱帯雨林、1854年から1862年までマレー諸島に昆虫採集の旅に出ました。ダーウィンの旅は、ウォレスが後にマレー諸島を旅したように、いくつかの島々を経由しており、最後に訪れたのはエクアドルの西にあるガラパゴス諸島でした。これらの島々でダーウィンは、異なる島々の生物の種を観察し、明らかに似ていながら、はっきりとした違いがあることを確認しました。例えば、ガラパゴス諸島に生息するフィンチは、独特の嘴の形をした数種類から構成されていました(図18.2)。ガラパゴス諸島に生息する種は、嘴の大きさや形が段階的に変化しており、最も似ている種の間の差は微々たるものです。彼は、これらのフィンチが南米本土の別のフィンチ種に酷似していることを観察しました。ダーウィンは、この島嶼種が本土の原種の一つから変化した種である可能性を考えました。さらに研究を進めると、フィンチの嘴の違いは、その鳥が特定の種類の餌を獲得するのに役立っていることがわかりました。例えば、種子を食べるフィンチは、種子を割るために強く太い嘴を持ち、昆虫を食べるフィンチは、獲物を刺すために槍のような嘴を持っていたのです。
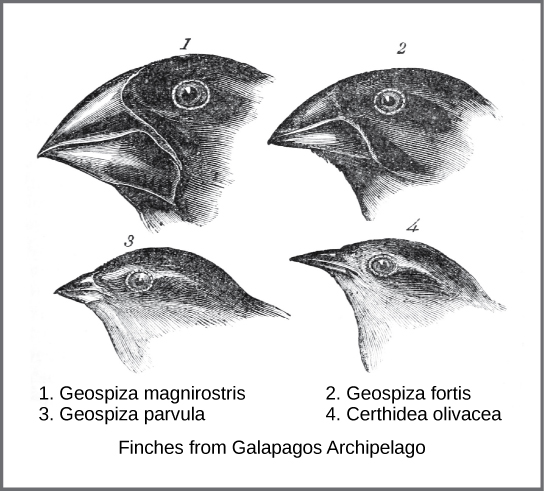
ウォレスとダーウィンは、他の生物にも同じようなパターンがあることを観察し、そのような変化がどのようにして起こるのか、なぜ起こるのかについて、同じ説明を独自に展開しました。ダーウィンはこの仕組みを自然選択と呼びました。自然選択とは、「適者生存」のことで、有利な特性を持つ個体がより多く繁殖し、その形質のために環境変化を生き抜くことをいいます。これが、進化的な変化をもたらすのです。
例えば、ダーウィンはガラパゴス諸島のゾウガメの個体群を観察し、乾燥した低地の他の島々に生息する個体よりも首が長いことを確認しました。これは、首の短いカメよりも、より多くの葉に手が届き、より多くの餌にありつけるからといえます。干ばつで葉が少なくなっても、より多くの葉に手が届くカメのほうが、手が届かないカメより食べて生き延びるチャンスがあったのです。その結果、首の長いカメは繁殖に成功しやすく、首の長い特性を子孫に受け継ぐことができました。そうしていくうちに、首の長いカメだけが集団で存在するようになるのです。
ダーウィンは、自然選択は自然界に存在する3つの原理の必然的な結果であると主張しました。1つ目は、生物の特性のほとんどは、親から子へと受け継がれるということ。ダーウィンやウォレスを含め、当時は誰もこの仕組みを知りませんでしたが、これは一般的な理解でした。2つ目は、子孫は生存能力よりも多く生産されるため、生存と生殖のための資源は限られているということ。すべての生物の生殖能力は、その数を支える資源の利用可能性を上回っています。そのため、各世代で資源をめぐる競争が起こるのです。ダーウィンもウォレスも、人間の個体数との関係でこの原理を説明した経済学者トマス・マルサス Thomas Malthus のエッセイを読んだことでこの原理を理解しました。そして3つ目は、子孫はその特性において互いに異なっており、その差異は遺伝するということです。ダーウィンとウォレスは、限られた資源を獲得するために最も有利な特性を持つ子孫は、競争力のない個体よりも生き残り、より多くの子孫を残すことができると考えたのです。特性は遺伝するので、その特性は次の世代でよりよく表現されることになります。これは、ダーウィンが「変化を伴う継承」と呼んだプロセスで、何世代にもわたって集団に変化をもたらすことになります。最終的には、自然選択によって、集団はその地域の環境により適応するようになります。自然選択は、適応進化をもたらす唯一のメカニズムとして知られています。
1858年、ダーウィンとウォレス(図18.3)は、ロンドンのリンネ協会で、自然選択の考え方を論じた論文を発表しました。そして翌年には、ダーウィンの著書『種の起源』が出版されました。この本には、自然選択による進化についての彼の主張がかなり詳細に概説されていました。

自然選択による進化の事例を記録し、発表することは困難であり、時間がかかります。ガラパゴスフィンチはその好例です。ピーター・グラント Peter Grant とローズマリー・グラント Rosemary Grant 夫妻は、1976年以来毎年ガラパゴスフィンチの個体群を調査し、自然選択の重要な証拠を示してきました。グラント夫妻は、ガラパゴス諸島のダフネ・マジョール島のガラパゴスフィンチで、くちばしの形の分布が世代間で変化していることを発見しました。この鳥は、広く深い嘴を持つ鳥と、薄い嘴を持つ鳥とで、嘴の形の変化を受け継いできたのです。エルニーニョの影響で平年より降水量が多かった時期には、大型の鳥が食べる大きな固い種子が不足し、小型の鳥が食べる小さな柔らかい種子が豊富にありました。そのため、小柄な鳥は生き残り、繁殖することができました。このエルニーニョの後の数年間、グラントらは集団のくちばしの大きさを測定し、平均的な嘴の大きさが小さくなっていることを発見しました。嘴の大きさは遺伝するため、嘴の小さい親はより多くの子孫を残し、嘴はより小さなサイズに進化していったのです。1987年に状況が改善され、より大きな種子が手に入るようになると、平均的な嘴のサイズが小さくなる傾向は収まりました。
Career Connection:フィールド生物学者
多くの人が、娯楽のためにハイキング、洞窟探検、スキューバダイビング、山登りなどに参加しています。また、野生動物との出会いを求めて参加される方も多いと思います。アウトドアを体験することは、とても楽しいことであり、元気を与えてくれるものです。もし、あなたの仕事が大自然の中で働くものだとしたらどうでしょう?フィールド生物学者は、屋外の “フィールド”で仕事をするのが基本です。この場合のフィールドとは、屋外のあらゆる場所を指し、水中であっても同様です。フィールド生物学者は、通常、特定の種、生物のグループ、あるいは単一の生息地を中心に研究を行います(図18.4)。

多くのフィールド生物学者が、未記録の新種を発見することを目的の一つとしています。このような発見は、自然界への理解を深めるだけでなく、医学や農業などの分野でも重要な技術革新につながります。特に植物や微生物の種は、新しい薬効や栄養の知識を明らかにすることができます。また、生態系において重要な役割を果たす生物や、希少な生物は保護が必要な場合もあります。これらの重要な生物種が発見されれば、研究者は環境規制や法律の根拠として利用することができるのです。
進化の過程とパターン
自然選択は、集団の中の個体間に変異、つまり違いがあって初めて発生するものです。重要なことは、これらの差異は何らかの遺伝的根拠を持っていなければならないということです。そうでなければ、自然選択は次の世代での変化につながりません。この点は非常に重要です。なぜなら、例えば、遺伝子の違いではなく、栄養状態が良いために身長が伸びるなど、非遺伝的な理由で個人差が生じることがあるからです。
集団における遺伝子の多様性は、主に突然変異と有性生殖の2つのメカニズムから生まれます。突然変異はDNAの変化であり、どの集団においても新しい対立遺伝子、すなわち新しい遺伝的変異を生み出す究極の源です。突然変異が引き起こす遺伝的変化は、表現型に対して3つの結果のうちの1つをもたらします。まず、突然変異は生物の表現型に影響を及ぼし、それにより生存の可能性が低くなったり、子孫の数が減ったりすると、適応度が低下します。あるいは、突然変異は、適応度に有益な影響を与える表現型を生み出すかもしれません。また、多くの突然変異は表現型の適合性に影響を及ぼしません。私たちはこれを中立的な突然変異と呼んでいます。突然変異はまた、その表現型に表れる生物の適応度に対して、小さな効果から大きな効果まで、あらゆる大きさの効果を持つ可能性があります。有性生殖もまた遺伝的多様性をもたらします。二人の親が繁殖するとき、対立遺伝子の特異的な組み合わせが集合して、それぞれの子孫に特異的な遺伝子型、ひいては表現型が生まれるのです。
生物が現在の環境で生存し、繁殖するために役立つ遺伝的形質を、適応と呼びます。科学者は、生物の集団が環境に適応することを、その集団の環境への適合度を高めたり維持したりする遺伝的変異が時間の経過とともに生じたとき、と表現しています。カモノハシの足には水かきがありますが、これは泳ぐための適応です。ユキヒョウの厚い毛皮は、寒さの中で生活するための適応、チーターの速いスピードは、獲物を捕らえるための適応です。
ある形質が好ましいかどうかは、現在の環境条件によって決まります。環境条件が変化することもあるので、同じ形質が常に選択されるとは限りません。例えば、湿潤な気候で育つ、水を節約する必要のない植物種を考えてみましょう。大きな葉は、太陽からより多くのエネルギーを得ることができるため、選択されました。大きな葉は、小さな葉よりも維持するために多くの水を必要とし、湿った環境は大きな葉を支えるために好条件を提供していました。しかし、数千年の時を経て、気候が変化し、水の余剰がなくなってしまいました。その結果、自然選択の方向が変わり、葉の小さな植物が選択されるようになりました。なぜなら、これらの個体群は、新しい環境条件を生き抜くために水を節約することができたからです。
種の進化は、形や機能に膨大な変異をもたらしています。時には、進化によって、互いにとてつもなく異なる生物群が生まれることもあります。ある共通点から多様な方向に進化した2つの種を分岐進化と呼びます。このような分岐進化は、顕花植物の生殖器官の形態に見ることができます。これらは基本的な解剖学的構造は同じですが、異なる物理的環境での選択と異なる種類の送粉者に対する適応の結果、まったく異なる姿になりました(図18.5)。

また、似たような表現型が遠縁の種で独立して進化するケースもあります。例えば、コウモリと昆虫の両方で飛行が進化し、どちらも私たちが翼と呼ぶ構造を持っていますが、これは飛行への適応といえます。しかし、コウモリの翼と昆虫の翅は、元々全く異なる構造から進化してきました。このように、共通の祖先を持たない種で、同じような形質が独立して進化する現象を収斂進化と呼びます。2つの種は、飛ぶという同じ機能にたどり着きましたが、互いに別々にそれを行ったのです。
このような物理的な変化は、膨大な時間をかけて起こるもので、進化がどのように起こるかを説明するのに役立ちます。自然選択は個々の生物に作用し、それが種全体を形成することができます。自然淘汰は個体には一世代で作用しますが、種全体の遺伝子型が進化するためには数千年、数百万年かかることもあります。このような長い時間をかけて、地球上の生物は変化してきたし、今も変化し続けているのです。
進化の証拠
進化の証拠は説得力があり、かつ広範です。生物学者は、生命システムのあらゆる階層の組織に、過去と現在における進化の痕跡を見て取ることができます。ダーウィンは、著書『種の起源』の大部分を、進化と一致する自然界のパターンを特定することに費やしましたが、ダーウィン以来、我々の理解はより明確かつ広範になっています。
化石
化石は、過去の生物が現在の生物と同じではないことを示す確かな証拠であり、進化の進行を示します。科学者たちは、世界中から集められた化石を年代別に分類し、その生物が相対的にいつ生きていたかを判断します。その結果得られる化石記録は、過去の物語を語るとともに、何百万年にもわたる形態の進化を示してくれます(図18.6)。例えば、科学者たちは、ヒトと馬の進化を示す非常に詳細な記録を復元しています(図18.6)。クジラのひれは、鳥類や哺乳類の付属肢と同じような形態をしており(図18.7)、これらの種が共通の祖先を持つことがわかります。

解剖学と発生学
進化を示すもう一つの証拠は、生物に同じ基本的構造が存在することです。たとえば、ヒト、イヌ、鳥、クジラの付属肢の骨は、共通の祖先の付属肢に由来するため、全体的に同じ構造をしています(図18.7)。時間の経過とともに、進化によって骨の形や大きさが種によって変わりましたが、全体としては同じレイアウトを保っているのです。科学者たちは、こうした意味での類似部分を相同的構造と呼んでいます。
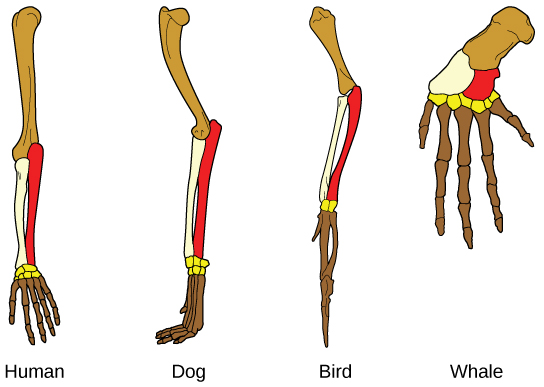
生物の中には、全く機能を持たず、過去の共通祖先の名残と思われる構造も存在します。このような、機能を持たない未使用の構造を、痕跡器官と呼びます。例えば、飛べない鳥の翼、一部のサボテンの葉、クジラの後脚の骨などです。もっとも、すべての類似性が相同的構造を表しているわけではありません。「進化的関係の決定」で説明しますが、類似した特徴が、進化上の密接な関係ではなく、環境の制約によって生じる場合、それは相似または同形形質とされます。例えば、昆虫はコウモリや鳥のように飛ぶために翅を使いますが、翅の構造も胚の起源も全く異なります。これらは相似的構造になります(図20.8)。
学習のためのリンク
人体の骨について探求する動画
もう一つの進化の証拠は、同じような環境を共有する生物において形態が収束することです。たとえば、北極圏に生息するホッキョクギツネやライチョウなどは、無関係な動物種ですが、雪や氷に溶け込むために、冬季に白い表現型が選択されてきました(図18.8)。こうした類似性は、共通の祖先がいるからではなく、同様の選択圧によって生じたものです。つまり、捕食者に見られないことで恩恵が得られたのです。

発生学は、生物が成体へと成長する過程を解剖学的に研究する学問ですが、現在では大きく異なる生物グループ間の関連性を示す証拠にもなっています。胚での突然変異は、成体においてより大きな影響を与えるため、胚の形成自体は保存される傾向にあります。その結果、あるグループには存在しない構造が、胚の形態ではしばしば現れ、成体または幼体の形態になると消えてしまう、ということが起こります。例えば、ヒトを含むすべての脊椎動物の胚では、発生初期のある時点で鰓裂と尾が現れます。これらは陸生群の成体では消失しますが、魚類や一部の両生類など水生群の成体では維持されます。ヒトを含む大型類人猿の胚は、発生過程で尾の構造を持ちますが、生まれたときには失われています。
生物地理学
地球上の生物の地理的分布のパターンは、地質学的時間における地殻変動と連動した進化によって最もよく説明できます。超大陸パンゲアが分裂する以前(約2億年前)に進化した大集団は、世界中に分布しています。また、ローラシア大陸から形成された北部大陸や、ゴンドワナ大陸から形成された南部大陸には、それぞれ固有の動植物が存在するなど、地球上の各地域でパンゲア分裂後に進化した集団が現れています。オーストラリア、アフリカ南部、南米には、南半球の超大陸ゴンドワナ大陸が分裂する以前に、ヤマモガシ科の植物が最も多く生息していました。
オーストラリアにおける有袋類の多様化と他の哺乳類の不在は、オーストラリアが長い間孤立していたことを反映しています。オーストラリアには、他では見られない固有種が数多く存在します。これは、海によって隔離され、種の移動ができない島々によく見られる現象です。これらの種は、時間とともに進化的に分岐し、本土に存在する祖先とはまったく異なる外見を持つ新しい種になります。オーストラリアの有袋類、ガラパゴス諸島のフィンチ、ハワイ諸島の多くの種は、その起源は一つでありながら、本土の祖先種とは遠く離れた関係を示しています。
分子生物学
解剖学的構造と同様に、生命の分子構造も、変化を伴う継承を反映しています。DNAの普遍性は、すべての生命に共通の祖先がいることを示す証拠です。遺伝情報、DNA複製、発現という生命の基本的な区分は、リボソームや膜構造など、他の保守的な構造における大きな構造上の違いに反映されています。一般に、生物集団の近縁性はDNA配列の類似性に反映されます。これは、まさに共通の祖先からの由来と多様化から予想されるパターンといえます。
また、DNA配列は、進化のメカニズムの一端を明らかにしました。例えば、タンパク質の新しい機能の進化は、遺伝子重複の後に起こることが多く、突然変異、選択、浮動(集団の遺伝子プールの偶然による変化)により、1つのコピーは自由に変化し、2つ目のコピーが機能するタンパク質を生成し続けられることが明らかになっています。
進化論に対する誤解
進化論は、ダーウィンが提唱した当初は賛否両論がありましたが、『種の起源』の出版後20年のうちに、特に若い生物学者を中心にほぼ全面的に受け入れられました。しかし、進化論は難しい概念であり、その仕組みについて多くの誤解があります。
学習のためのリンク
このサイトでは、進化論にまつわる主な誤解を取り上げています。
”進化は単なる理論に過ぎない”
進化論の批判者は、「理論」という言葉の日常的な用法と科学者の用法を意図的に混同して、その重要性を否定します。科学では、「理論」とは、自然界の一連の観察に対して、徹底的にテストされ検証された説明の集合体であると理解されています。科学者は、原子、重力、そして相対性理論などの理論を持っており、それぞれが世界について理解された事実を説明しています。同じように、進化論は生物界に関する事実を記述しています。このように、科学における理論は、科学者による、その信用を失墜させようとする多大な努力に耐えてきたのです。これに対して、一般的な言葉での「理論」は、推測や提案された説明を意味する言葉です。この意味は、科学の概念における「仮説」に近いものです。進化論を批判する人が「単なる理論だ」と言うのは、それを裏付ける証拠がほとんどなく、まだ厳密な検証の段階にあることを暗に示しています。そしてこれは誤った認識なのです。
”個体が進化する”
進化とは、ある集団の遺伝的構成が時間とともに、具体的には世代を超えて変化することであり、特定の対立遺伝子を持つ個体が繁殖することによって生じるものです。個体が生涯の中で変化することは確かにあります。しかし、これは発達であり、個体が生まれながらにして獲得した一連の遺伝子によってプログラムされ、個体の環境と協調して変化するものです。ある特性の進化を考える場合、その形質の母集団における平均値の時間的な変化を考えるのがよいでしょう。例えば、ガラパゴス諸島のフィンチの嘴の大きさが自然選択によって変化しても、それは個々のフィンチの嘴が変化していることを意味するものではありません。ある時期に集団内の全個体の平均的な嘴の大きさを測定し、数年後に集団内で測定すると、進化の結果、この平均値は異なるものになります。ここで、1回目の測定から2回目まで生き残る個体もいるかもしれませんが、その個体の嘴の大きさは変わらないでしょう。しかし、そこには新しい個体が多く存在し、それが平均的な嘴の大きさの変化に寄与するのです。
”進化論は生命の起源を説明する”
進化論には生命の起源の説明が含まれているというのは、よくある誤解です。逆に、この理論の批判者の中には、生命の起源を説明することはできないと考える人もいます。進化論は生命の起源を説明しようとはしていません。進化論は、個体群が時間とともにどのように変化し、生命がどのように種の起源を多様化させるかを説明するものです。しかし、生命を定義する最初の細胞の起源のような、生命の始まりを明らかにするものではありません。重要なのは、生物学者が、地球上の生命の存在は、地球上の生命につながる出来事が繰り返される可能性を排除すると考えていることです。なぜなら、生命につながるような中間段階は、直ちに既存の生物の食料となるからです。
とはいえ、細胞や細胞以前の物体の中にDNAのような分子の形で遺伝の仕組みができてしまえば、その存在は自然選択の原理に従うことになります。より効果的な繁殖者が頻度を増やし、非効率的な繁殖者を犠牲にすることになるのです。進化論は生命の起源を説明するものではありませんが、生物以前の物体がある特性を獲得するまでの過程について、何かしらの示唆を与えてくれるかもしれません。
”生物は目的を持って進化している”
「生物は環境の変化に応じて進化する」というような表現はよくありますが、このような表現には2つの誤解があります。第一に、「生物は個々に進化する」と解釈してはなりません。この文は、「環境の変化に対応して集団が進化する」ということを縮めた表現です。しかし、この文を、進化が何らかの意図的なものであると解釈すると、第二の誤解が生じる可能性があります。環境が変化すると、集団の中のある個体、つまり特定の表現型を持つ個体が恩恵を受け、その結果、他の表現型に比べて割合的に多くの子孫を残すことになります。この結果、もしその特性が遺伝的に決定されているのであれば、集団に変化が生じます。
また、自然選択が働く変異は、すでに集団の中にあるもので、環境変化に応じて生じるものではないことを理解することが重要です。例えば、あるバクテリアの集団に抗生物質を投与すると、時間の経過とともに、抗生物質に耐性を持つバクテリアの集団が選択されることになります。遺伝子が引き起こす耐性は、抗生物質を塗ったから突然変異で生じたのではありません。耐性の遺伝子は、バクテリアの遺伝子プールの中に、おそらく低い頻度ですでに存在していたのです。抗生物質は、耐性遺伝子を持たないバクテリアの細胞を殺すので、耐性を持つ個体が強く選択されることになります。しかし、抗生物質に対する耐性変異は、抗生物質によって生じるものではないことが、実験によって明らかにされています。
より広い意味では、進化には目標がありません。種は時間とともに「より良く」なるのではないのです。種は単に、その時々の特定の環境において繁殖を最大化するような適応をもって、環境の変化に追従しているだけです。進化には、より速く、より大きく、より複雑な、あるいはより賢い種を作るという目標はないのです。ある種の中で進化する特性は、存在する変異と環境の関数であり、両者は常に非一様に変化しています。ある時期にある環境に適合した形質が、将来のある時期には致命的なものになる可能性も十分にあります。このことは、昆虫の種でも人間の種でも同じように言えることです。
📖おすすめ書籍
進化とは何か:ドーキンス博士の特別講義
『利己的な遺伝子』で世界的に有名な進化生物学者リチャード・ドーキンスが、一般人に向けて行った進化についての科学講演を書籍化したもの。親しみやすい語り口と視覚的にわかりやすい実験を通して進化とは何かについて解き明かす。
利己的な遺伝子 40周年記念版
進化生物学の古典的名著。本書を読んで進化論の提示する生命観を味わえば、明日からの世界がまったく違って見えてくるだろう。2017年に、一般投票による「英国史上最も刺激的な科学書」の第1位にも選ばれた。


コメント
[…] 18.1 進化とは […]
[…] 18.1 進化とは学習目標現在の進化論がどのように発展してきたかを説… […]